
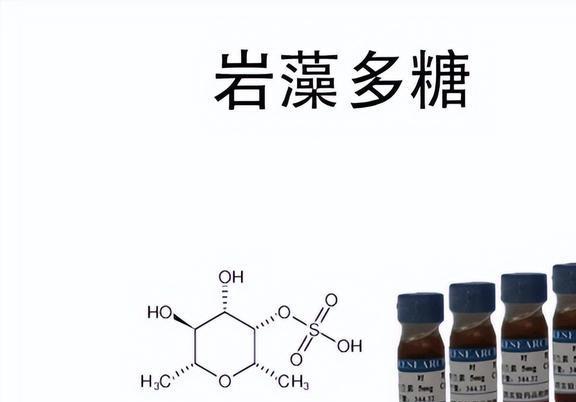
原来多糖是天然高分子化合物,免疫调节被认为是最重要的生物活性
多糖是一种由10个及以上的单糖聚合而成的天然高分子化合物,广泛存在于动植物中和微生物中,具有多种生物活性,比如调节免疫、抗肿瘤、抗氧化和降血糖等生理功能。
其中,免疫调节作用被认为是其最重要的一种生物活性。
研究人员从组织器官水平、细胞水平、分子水平以及肠道菌群出发,深入研究多糖免疫调节作用及其机制,证明了多糖免疫调节作用并非单一环节,而是多环节、多靶点调节机体的免疫功能。
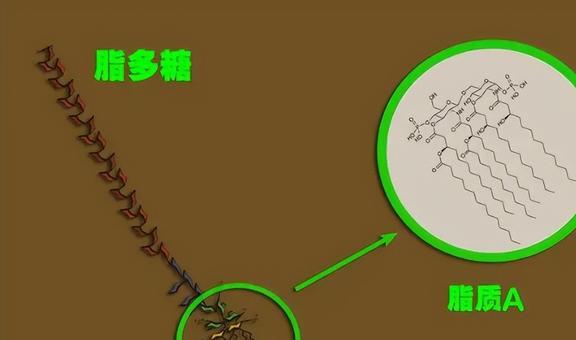
相比多糖的药效学研究,其药动学的研究进展较为缓慢。
由于多糖的相对分子质量较大且无荧光基团或紫外吸收,使得多糖的示踪和检测成为难点。
本文对近年来多糖体内药动学研究、免疫调节作用机制研究的技术进行总结,以期为多糖体内过程及其免疫调节机制研究提供一定的技术参考。

口服给予多糖后,部分多糖难以被唾液或胃液中的酶消化。
肠道中寄生着丰富的肠道菌群,以拟杆菌门和厚壁菌门为主,也包括少量放线菌门和变形菌门。
这些肠道微生物能够分泌多糖降解酶降解多糖,可促进机体对多糖的消化和吸收。
此外,小肠肠黏膜表面积较大,富含丰富的小肠绒毛和肠毛细血管,并且存在特殊的转运体,这些结构特征有助于多糖的体内吸收。
因此,多糖主要是通过肠道吸收后进入体内并发挥药理活性。
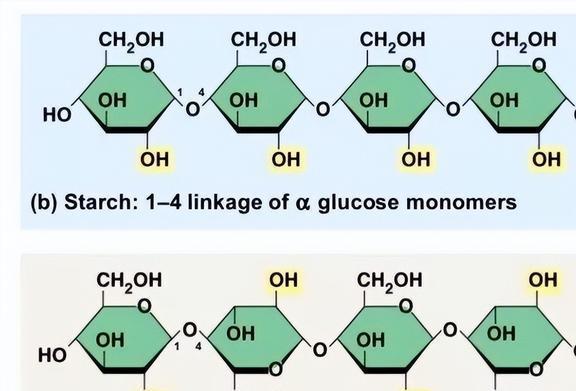
多糖在肠道的吸收方式主要有:细胞旁路通道转运、跨细胞途径和M细胞介导的转运。
多糖通过细胞旁路吸收是一个被动转运的过程,并且由肠上皮细胞紧密连接(tightjunction,TJ)控制,肠道不同节段的上皮细胞紧密连接的空隙各不相同,当肠上皮细胞紧密连接完全打开时,大分子量多糖即可吸收。
多糖的跨细胞途径吸收主要指多糖在肠细胞顶膜处通过胞吞作用从肠腔转运至细胞外液。
M细胞在内吞吸收多糖作用中非常活跃,与肠上皮细胞不同,M细胞的顶端膜缺乏正常的微绒毛,呈现出褶状结构。
此外,其基底膜深凹呈囊状,一些肠道相关的淋巴细胞聚集在此处形成派尔集合淋巴结,且M细胞表面含有多种多糖的特异性受体,可识别外源性多糖并运输给派尔集合淋巴结中的淋巴细胞。
因此,M细胞介导的转运途径对多糖的肠道吸收具有十分重要的意义。
随着研究的不断深入,多糖免疫调节机制的研究已经达到分子和受体的水平。
Toll样受体、清道夫受体、补体受体3和Dectin-1等是目前研究发现的主要多糖受体。

多糖通过识别并结合免疫细胞表面的特定受体,激活细胞内下游信号通路,进而促进相关细胞因子的表达,从而启动免疫反应并发挥免疫调节作用。
多糖与Toll样受体结合后,通过激活髓样分化蛋白88(MyD88)来促进核转录因子-κB(NF-κB)和丝裂原活化蛋白激酶(MAPK)分泌促进肿瘤坏死因子-α (TNF-α)、白细胞介素-1β(IL-1β)、白细胞介素-6(IL-6)等促炎细胞因子。
多糖和清道夫受体结合后,激活p38丝裂原活化蛋白激酶(MAPKp38)及NF-κB两条信号通路,继而促进TNF-α和白细胞介素-12(IL-12)的分泌。
补体受体3(CR3)能识别β-葡聚糖,与白细胞分化抗原14(CD14)形成复合体,共同激活磷脂酶(PLC),继而激活蛋白激酶(PKC)和磷脂酰肌醇-3-激酶(PI3-K),通过MAPK或NF-κB信号通路调控相关细胞因子的表达。
Dectin-1与β-葡聚糖特异性结合后,激活细胞内酪氨酸激酶(Syk),进而激活MAPK或NF-κB信号通路并释放细胞因子。
此外,dectin-1能增强Toll样受体介导的NF-κB信号通路,共同发挥作用,引起免疫应答。
正常情况下,肠道菌群与机体免疫调节系统处于动态平衡状态,维持着肠道微生态的稳定。
多糖进入肠道后与肠道菌群发生相互作用,这些微生物群介导的生物转化将大分子多糖降解为低聚糖或单糖,同时,多糖也能够调节肠道菌群相对丰度,多糖被分解后作用于肠系膜淋巴系统、派尔集合淋巴结以及肠上皮内淋巴细胞,从而下调TGF-β、干扰素-γ (INF-γ)、IL-1β和IL-6等炎症因子来减轻炎症反应。
多糖的免疫调节活性与肠道菌群的作用密切相关。
多糖进入肠道后在肠道菌群的作用下降解为单糖或低聚糖,经代谢最终生成一系列短链脂肪酸(SCFAs)。
而SCFAs对调控宿主免疫具有重要作用。
SCFAs主要通过激活免疫细胞趋化、分化、增殖和凋亡来参与机体免疫调控,这些调控过程主要通过激活G蛋白偶联受体(GPCRs)和抑制组蛋白去乙酰化酶(HDAC)活性来实现。

SCFAs与G蛋白偶联受体109a(GPR109a)结合,抑制NF-κB的激活,从而下调TNF-α、白细胞介素-2(IL-2)、IL-6和转化生长因子-β (TGF-β)等炎症因子的表达,GPR109a的激活还可促进Treg细胞的转录因子Foxp3的表达,使得Treg细胞数目增加,从而抑制炎症的发生。
SCFAs通过激活GPR41促进缺氧诱导因子1α (HIF-1α)的表达来上调抗炎因子白细胞介素-22(IL-22)的产生,提高肠道细胞的免疫活性。
G蛋白偶联受体43(GPR43)的激活可促进Th1细胞的分化以及炎性因子白细胞介素-10(IL-10)的释放。
此外,SCFAs可直接通过抑制HDAC活性来调节免疫反应,当HDAC受到抑制后会导致NF-κB失活,从而减少促炎细胞因子的生成。
SCFAs能够刺激肠黏膜中B细胞分泌免疫球蛋白A(IgA),并且其诱导分泌型免疫球蛋白A(sIgA)的释放,维持机体免疫平衡。

多糖能够与细胞膜表面的分子结合形成糖蛋白,进而参与信号传导并发挥免疫调节作用。
然而由于多糖本身无发光基团,导致其难以在复杂体系中定性或定量分析,进而使得多糖在体内的吸收、分布、代谢和消除的相关研究存在相对不足,这在一定程度上限制了多糖药代动力学的深入研究。
目前,多糖体内药动学研究以荧光标记法和同位素标记法较为常用。
荧光标记法是将多糖分子上的活性基团(如羟基、氨基、羧基等)与荧光素以共价偶联的方式进行结合,通过检测荧光强度对多糖进行定性、定量分析。
被标记后的多糖因具有荧光发光基团,可排除内源性多糖物质的干扰。
研究人员通过该技术实现对多糖的吸收、分布、代谢、排泄各环节的监测,有利于多糖药代动力学的深入研究。
Liang等采用荧光素5-异硫氰酸酯(FITC)标记半乳糖基化壳聚糖(GC)的方法研究其在小鼠体内的器官靶向性,结果表明FITC-GC在静脉注射5min后在肝脏显著富集,48h后仍可在肝脏中检测到。
相比之下,FITC-GC在肝脏中比FITC-CS(未加半乳糖修饰的壳聚糖)富集浓度更高。
Kaneo等研究了静脉注射后大鼠血浆中FITC标记的阿拉伯半乳聚糖(FA)的全身分布与消除,研究发现,FA在静脉注射30min后在血浆中被清除,随后主要分布在肝脏和肾脏中。

Zhang等研究了经FITC标记的当归多糖(ASP)静脉给药后的药代动力学信息,研究发现,ASP在静脉注射后能够迅速从血液中消除并以高特异性分布到肝脏,且在体内未发生降解。
荧光标记法不仅可以给多糖定量,还可以确定多糖在体内是否被降解,这是荧光标记法的优点。
然而,荧光标记法在多糖药代动力学研究中仍存在一定的不足,荧光标记后的多糖的敏感性不强,仅能在浓集器官和血液中检测到,因此会增加操作的复杂性。
FITC中的异硫氰酸酯基团与氨基具有高反应性,在一定条件下可直接发生亲核加成反应,因此,FITC被广泛用于多糖的荧光标记。
而羟基对N-甲基靛红酸酐(MIA)的反应基团苯并噁嗪环具有特异性反应,直接以偶联的方式结合。
含糖醛酸的酸性多糖可用含氨基的荧光素ANDSA直接标记,用过量的1-乙基-3-(3-二甲基氨丙基)-碳化二亚胺(EDC)将多糖活化后,直接加入荧光素ANDSA接入发光基团。
因此,不同多糖荧光标记的位置也各不相同,常用的荧光标记位点主要分为三类,

Group
Fluorescein
Polysaccharide
Hydroxylgroupsofpolysaccharides(-OH)
5-DTAF
Konjacmannan,yeastpolysaccharides
FITC
Angelicapolysaccharides,Ganodermalucidumpolysaccharides,Gojiberrypolysaccharides
RBITC
Dendroglossanpolysaccharides,Dextran
MIA
Dextran
Aminogroupsofpolysaccharides(-NH2)
AF
Seaweedpolysaccharides
Aldehydegroupofpolysaccharides(-CHO)
APTS
Malto-oligosaccharides
ANTS
Sulfatephycosan
放射性同位素由于其放射强度和半衰期各不相同,在体内的药物代谢动力学研究会呈现不同的特点。
为了解决多糖体内检测技术的局限带来的一系列难题,放射性同位素标记技术也开始运用到多糖的代谢动力学研究中,由于放射性同位素能够自发地发射特征谱线,在体外与多糖分子结合后可以利用核探测器、电子计算机断层扫描(computedtomography,CT)对多糖进行定量、定位检测,用于追踪多糖在体内发挥免疫调节作用的代谢途径。
常用的放射性同位素有:14C、3H、125I、99mTc等。
相关文章
-

陈都灵一袭绿色抹胸长裙 站在柳树下宛若自然精灵
近日,陈都灵最新造型美照释出,她身着一袭绿色抹胸长裙,站在柳树下宛若自然精灵,神秘而又清新。...
2025-04-24 09:21:51 都灵站在抹胸 -

梁朝伟刘嘉玲周润发齐聚 为杜琪峰庆生掀起回忆杀
4月22日晚,刘嘉玲于社交账号上发布了与梁朝伟、周润发、张叔平等好友为杜琪峰庆生的合影,画面中众人笑容满面,多年情谊尽显。刘嘉玲周润发同框合影,刘嘉玲微卷的利落短发造型十分吸睛。...
2025-04-24 09:21:44 刘嘉玲梁朝伟杜琪 -

林心如就黄子佼事件强硬表态:对虐待及影片持有行为零容忍
台媒消息显示,林心如于近期就黄子佼藏匿未成年人性影像一事再度发声。她郑重表明立场,明确表示对虐待未成年人以及持有相关影像的行为绝无妥协余地。...
2025-04-24 09:21:41 强硬影片行为 -

-

董璇泳池旁喝椰子水好惬意 穿优雅礼裙身姿曼妙
近日,董璇在社交平台上晒出一组写真美照,并配文称:“提前在广州感受了夏日赛道。”照片中,她身着一袭优雅礼裙在游泳池旁喝椰子水,面露明媚笑容,十分惬意。...
2025-04-24 09:21:14 椰子泳池曼妙 -

-

形势比人强,特朗普关税战打不下去了!
原创: 后沙来源微信公众号:后沙已获转载授权昨天在短短不到6个小时之内,特朗普发动的关税战又变调了。北京时间0时左右,美国财政部长贝森特在摩根大通的闭门会议上,放风称,中美之间的高关税是不可持续的,预计会降温。预计什么,预料到中国的态度?自己信吗?1点左右,白宫发言人莱维特在新闻发布会上也表达了类似的态度。5点左右,特朗普亲自确认,针对中国进口商品的关税税率不会维持在当前水平(145%),“它会大幅下降,但也不会降至零,中国将对最终的关税税率非常满意”。特朗普还说了些肉麻的话,要和中国幸福地生活在一起(v...
2025-04-24 09:21:03 关税不下去了形势 -

-

怂是真认怂,事是真没完!
作者:码头整薯条的海鸥来源:码薯学人今天我们来聊聊服软的特朗普。在美国时间4月22号发生了4件事:特朗普公开说开始考虑降低对华关税税率(强调不是0);特朗普放出消息说中美贸易谈判进展顺利;特朗普表示无意解雇美联储主席鲍威尔,但是还是要求美联储降息;特朗普的财长贝森特在22号摩根大通的闭门投资会上明确表示中美贸易战不能长期持续。这4条消息合在一起,我们可以明确一点,特朗普在释放善意。但是,善意不代表胜利。果然,就在4月23日,美国媒体突然传出消息说中美芬太尼谈判陷入僵局。22号的宣传口径表明特朗普确实在认怂...
2025-04-24 09:21:01 怂是真认怂事是真 -

特朗普不想打关税战了,想和中国幸福地生活在一起!
原创: 一棵青木来源公众号:远方青木已获转载授权特朗普发动的关税战又进入新阶段了,以一种你想都不敢想的方式。4月23日,特朗普公开发言称对中国的145%关税太高了,自己很快会大幅度降低,但不会降低到零。特朗普还说,中国将对最终的关税税率非常满意,但他们必须跟美国达成协议。还有更离谱的,特朗普说:“我认为中国会很高兴,我们会幸福地生活在一起”。没错,真的是这么说的。中文翻译看起来非常肉麻,但原文其实更肉麻,特朗普使用的英文单词原文是“very happily”。不仅要在一起(live together),还...
2025-04-24 09:20:59 关税中国幸福



